Cambiums et arborescences
Toute plante croît d’abord en longueur au niveau caulinaire et racinaire, via la prolifération de cellules ou de territoires cellulaires apicaux à propriétés embryonnaires que l’on appelle méristèmes et qui produisent des tissus dits primaires, avec pour conséquence que les tiges et les racines produites ont un diamètre défini et constant. Or d’un point de vue biomécanique, une tige sera d’autant plus grande ou plus haute (maximisant ainsi la dispersion des spores) que son diamètre est grand, et un diamètre limité n’autorise alors qu’une hauteur limitée, quelle que soit la densité des tissus de soutien. Une tige peut ainsi augmenter sa croissance verticale si elle peut également croître en épaisseur. Dans de nombreux groupes, cette croissance en épaisseur pourra se réaliser via la mise en place de méristèmes dits secondaires. Au niveau caulinaire, un méristème se met en place sous l’épiderme, c’est le phellogène ou assise subéro-phellodermique qui produira l’écorce ou périderme à fonction protectrice (dans la racine, il prend naissance dans le péricycle sous l’endoderme). Dans la tige et la racine, entre le xylème et le phloème, un méristème distinct, le cambium ou assise libéro-ligneuse, produira vers l’intérieur le xylème secondaire ou bois et vers l’extérieur le phloème secondaire ou liber. La production conjointe de bois et de liber définit le cambium bifacial et caractérise les Lignophytes (du liber était peut-être également produit par convergence chez les Sphenophyllales (Equisetidae) éteintes, mais c’est encore discuté). Chez les autres formes arborescentes non Spermatophytes, abondantes au Paléozoïque et actuellement éteintes (comme les Lepidodendron – Lycopodiidae, et les Calamites - Equisetidae), le cambium était unifacial et ne produisait que du bois. L’acquisition d’un tissu fortement lignifié au centre de la tige (le cylindre de bois) et l’accroissement du diamètre autoriseront donc l’apparition de plantes de très grandes tailles, mesurant plusieurs dizaines de mètres de haut, voire dépassant les 100 mètres pour certains séquoias (Pinidae) et eucalyptus (Magnoliidae) actuels. On notera cependant que chez les Lepidodendron et formes arborescentes apparentées du Paléozoïque, le cylindre de bois était relativement peu épais et que l’essentiel du soutien était assuré par une écorce très développée. L’arborescence, et donc l’acquisition de cambiums, est apparue par convergence de nombreuses fois entre le Dévonien moyen et le Dévonien terminal, chez les « cladoxylopsides », chez les Equisetidae, chez les Lycopodiidae et chez les Lignophytes et vraisemblablement dès le Carbonifère chez des groupes éteints de fougères. C’est d’ailleurs au Dévonien supérieur que vont se développer les premiers vrais écosystèmes forestiers constitués essentiellement de « progymnospermes » dont les Archaeopteridales (voir chapitre sur les Lignophytes) à l’ombre desquelles vont se différencier les premiers Spermatophytes, les premières fougères et les premiers animaux tétrapodes. Actuellement l’arborescence et les vrais troncs (avec du bois) ne s’observent plus que chez les Spermatophytes ligneux. L’arborescence sans bois sera aussi expérimentée dans certains groupes, chez des fougères arborescentes (certaines Marattiales éteintes du Carbonifère comme les Psaronius, les Cyatheales actuelles et d’autres groupes dont certains Blechnum tropicaux) et chez les Lilianae (qui n’ont plus de cambium fonctionnel) comme les palmiers (Arecaceae), les Pandanus, et de nombreuses Asparagales tropicales. Dans ce type d’arborescence, l’absence d’un cylindre de bois est plus ou moins compensée par la prolifération de tissus sclérifiés, voire par de l’épaississement de tissus primaires (chez les palmiers), mais le diamètre restant limité, les tailles n’atteignent jamais celles des formes ligneuses.
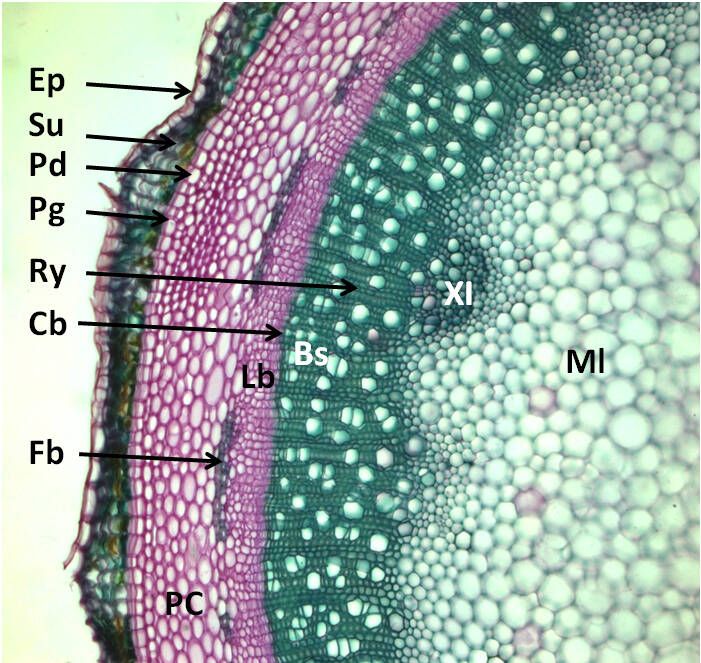
Tissus secondaires chez les Lignophytes.
Coupe transversale dans une tige âgée de Sureau (Sambucus nigra, Adoxaceae), coloration carmin-vert d’iode, le carmin colore la cellulose en rose, le vert d’iode la lignine (et la subérine qui imprègne l’écorce) en bleu-vert ; Ml = moelle ou parenchyme médullaire, XI = xylème primaire, Bs = bois ou xylème secondaire, Ry = rayon du bois, Cb = cambium (ici bifacial), Lb = liber ou phloème secondaire, Fb = fibres périphloémiennes (sclérenchyme, qui ici remplace le phloème primaire qui est écrasé par le développement des tissus secondaires), PC = parenchyme cortical, Pd = phelloderme, Pg = phellogène, Su = suber, Ep = épiderme qui se desquame ; Phelloderme, phellogène et suber forment le périderme ou écorce. Le bois est constitué d’éléments dits verticaux (ou longitudinaux) qui sont les trachéides et/ou vaisseaux, des fibres et parfois du parenchyme vertical, et des éléments horizontaux, des rayons de parenchyme orientés radialement. Le tout est disposé radialement (le cambium produit des cellules alignées radialement) et est (normalement) lignifié.
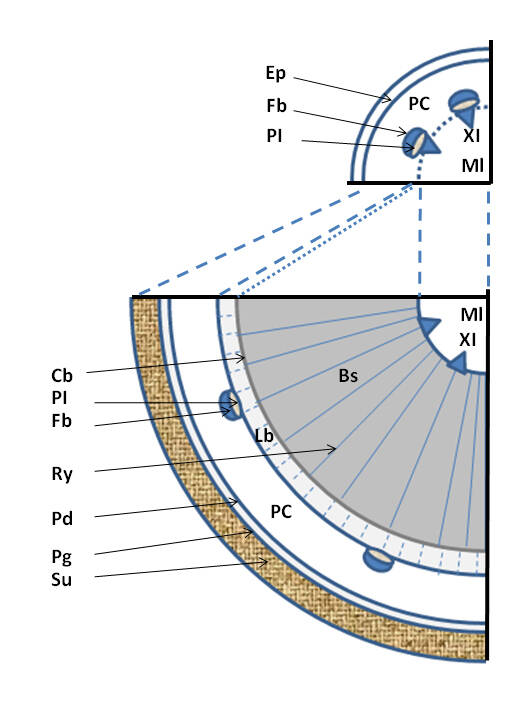
Mise en place des tissus secondaires chez les Lignophytes.
(Haut) Représentation schématique de l’anatomie d’une tige jeune sans tissus secondaire, le cercle en pointillé indique la position théorique du cambium, (Bas) représentation schématique de l’anatomie de la même tige âgée et épaissie via la mise en place des tissus secondaires ; Ml = moelle ou parenchyme médullaire, XI = xylème primaire, Bs = bois ou xylème secondaire, Ry = rayon du bois, Cb = cambium (ici bifacial), Lb = liber ou phloème secondaire, PI = phloème primaire, Fb = fibres périphloémiennes (sclérenchyme), PC = parenchyme cortical, Pd = phelloderme, Pg = phellogène, Su = suber, Ep = épiderme ; Phelloderme, phellogène et suber forment le périderme ou écorce (ici l’épiderme a disparu avec le développement de l’écorce).
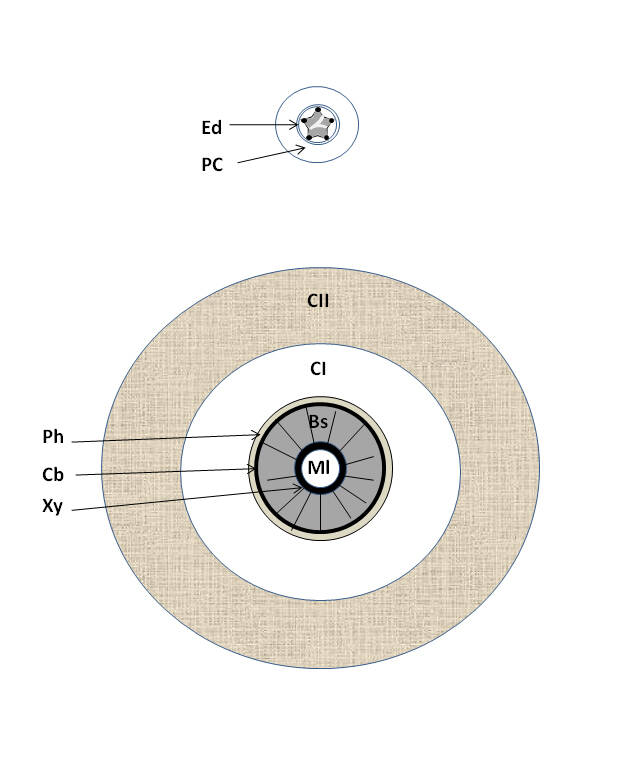
Mise en place des tissus secondaires chez les Lycophytes arborescents (éteints).
(Haut) Représentation schématique de l’anatomie d’une tige jeune de Lycophyte sans tissus secondaire avec actinostèle typique, (Bas) représentation schématique de l’anatomie d’une tige de Lepidodendron arborescent éteint (voir chapitre Lycophytes) ; Ml = moelle ou parenchyme médullaire, Xy = xylème primaire, Bs = bois ou xylème secondaire, Cb = cambium (ici unifacial), PI = phloème primaire, PC = parenchyme cortical, CI = parenchyme cortical primaire, CII = parenchyme cortical secondaire (écorce secondaire) ; dans les formes arborescentes, les parenchymes corticaux ou cortex primaire et secondaire peuvent se subdiviser en diverses zones (non détaillées ici), la position du phellogène au sein du cortex secondaire n’est pas toujours identifiable ou supposée proche de la périphérie de la tige, la partie interne du cortex secondaire étant donc bien plus épaisse que son analogue (le phelloderme) du périderme des Lignophytes. Les formes jeunes des Lepidodendron sont protostéliques et la stèle va se médullariser avec l’âge, ce qui explique la présence d’une moelle de plus en plus large au fur et à mesure que l’on s’élève dans le tronc des formes adultes. Comme pour les Lignophytes, les tissus secondaires sont orientés radialement.

Créez votre propre site internet avec Webador