Diversité et évolution morphologique
La morphologie des premiers Embryophytes
Karl Niklas dans son ouvrage Evolutionary Biology of Plants publié en 1997 s’est intéressé à l’évolution de la morphologie chez les plantes terrestres via une approche modélisatrice qu’il confronte aux observations sur les fossiles et les actuels. En milieu aquatique, à partir d’un organisme unicellulaire originel, les organismes chlorophylliens pluricellulaires doivent développer une morphologie qui doit répondre à deux fonctions : la capture de la lumière pour la photosynthèse et les échanges gazeux pour la respiration. Niklas montre donc que les morphologies attendues efficaces vont du filament plus ou moins ramifié à la lame aplatie, elle-même pouvant être plus ou moins ramifiée, soit toutes les morphologies effectivement observables chez les « algues » marines et d’eau douce. Hors de l’eau, l’organisme est soumis à d’autres contraintes, en dehors du problème de dessiccation que les plantes terrestres vont prévenir avec la cuticule (absente chez les « algues »). Deux contraintes ou forces évolutives a priori antinomiques vont intervenir. La gravité va « maintenir plaqués » au sol les organismes, mais ces derniers pour optimiser la dispersion des spores doivent les produire le plus haut possible. Deux morphologies sont donc possibles et attendues, la lame aplatie plus ou moins ramifiée qui va permettre la colonisation du substrat mais qui est peu efficace pour la dispersion des spores ou le cylindre dressé qui est biomécaniquement la morphologie optimale pour croître verticalement et disperser des spores en hauteur mais peu efficace pour l’exploration du substrat. Les premiers Embryophytes vont combiner les deux, une forme végétative thalloïde (ou filamenteuse) pouvant se ramifier pour explorer le substrat horizontalement (le gamétophyte) et une forme cylindrique dressée portant un sporange (le sporophyte ou sporogone) pour la dispersion. Chez les formes à sporogone réduit (chez les Marchantiales par exemple), la génération sporophytique est amenée en hauteur par un archégoniophore gamétophytique cylindrique. Chez les Bryidae (ou mousses vraies), le gamétophyte devient lui-même cylindrique, ce qui permet, chez les formes acrocarpes en particulier, d’avoir une croissance verticale qui s’ajoute à celle du sporogone. On notera ici que la sélection de la croissance verticale dominante chez les plantes actuelles est liée initialement à la dispersion, avant d’être utilisée en contexte de compétition pour accéder à la lumière.
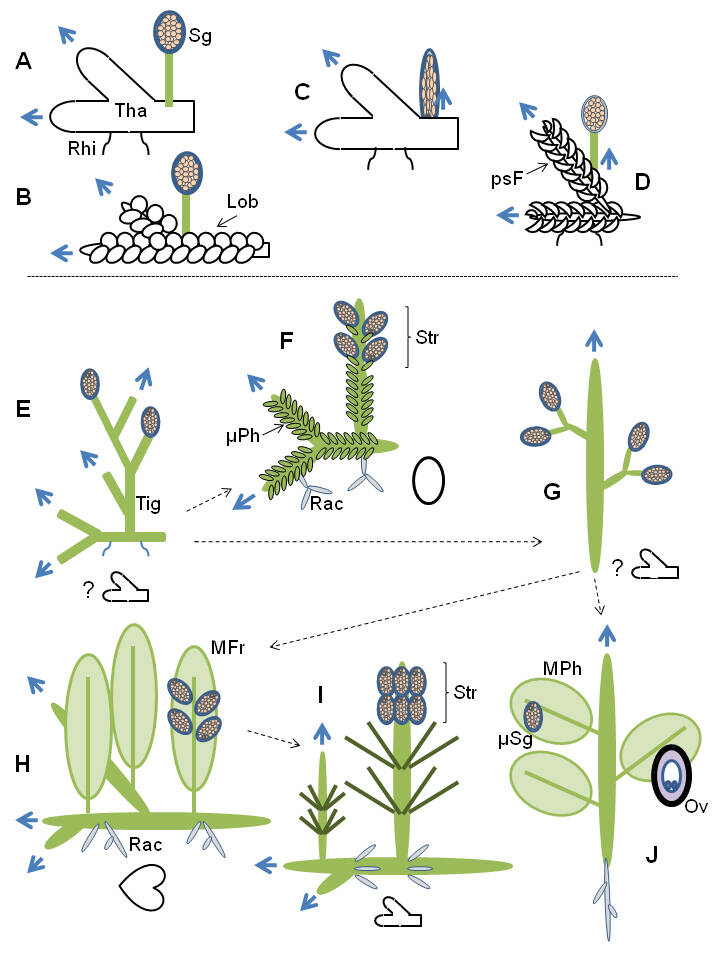
Diversité et évolution de la morphologie chez les Embryophytes.
Le gamétophyte est en blanc (certains marqués par un ? sont encore inconnus ou hypothétiques), le sporophyte est en vert, les flèches pleines indiquent le sens de la croissance des axes principaux, les flèches en pointillé les voies évolutives possibles ou inférées, les modalités de la reproduction sont détaillées dans le chapitre reproduction et dans les chapitres correspondant aux taxons ; A. Marchantiidae à thalle, B. Marchantiidae à lobes, C. Anthocerotidae (le sporophyte croît par la base), D. Bryidae, E. premiers polysporangiophytes (éteints), F. Lycopodiidae, G. premiers Euphyllophytes (éteints), H. Polypodiidae, I. Equisetidae, J. Spermatophytes ; Tha = thalle, Rhi = rhizoïdes, Lob = lobes, Sg = sporange, psF = pseudo-feuilles, Tig = tige (cormus), Rac = racines, µPh = microphylle, MPh = mégaphylle, MFr = mégafronde, Str = strobile, µSg = microsporange, Ov = ovule.
Diversité et évolution morphologique
Comme le montre également Niklas, la morphologie des plantes terrestres à sporophyte dominant doit répondre à trois contraintes et fonctions principales : la capture de la lumière, la dispersion des spores et le soutien. La capture de la lumière favorise la sélection d’organes à croissance horizontale. La dispersion des spores favorise la croissance verticale. Le soutien est nécessaire pour à la fois assurer une croissance verticale et porter les organes assimilateurs qui auraient tendance à se développer horizontalement. Chez les Bryidae, la capture de la lumière est assurée en grande partie par les pseudo-feuilles du gamétophyte. Chez les premiers Polysporangiophytes, le sporophyte dominant est réduit à une tige (cormus) ramifiée qui porte les sporanges. Certains axes sont horizontaux en assurant colonisation et exploration du substrat (il n’y a pas de système racinaire et l’absorption de l’eau et des minéraux est assurée par des petits rhizoïdes qui émergent directement des tiges) et les autres axes sont verticaux pour amener et soutenir les sporanges en hauteur. Les tiges sont aphylles et assurent autant la photosynthèse, que l’exploration du substrat et la production et le soutien des sporanges. Sur des axes verticaux, la photosynthèse est possible mais moins efficace que via des surfaces horizontales. La sélection d’organes horizontaux spécialisés apporterait un avantage certain en permettant de dissocier la fonction de soutien des sporanges de celle de la capture de la lumière, voire les compléter si les organes assimilateurs produits en hauteur portent également les sporanges. Cette dissociation apparait en premier chez les Lycophytes au Dévonien inférieur avec l’acquisition de la microphylle qui serait une émergence épidermique (énation) secondairement vascularisée (voir chapitre correspondant). Des rameaux latéraux spécialisés vont ensuite apparaitre indépendamment chez les Euphyllophytes via la croissance pseudo-monopodiale également au Dévonien inférieur (voir chapitre correspondant). Ces rameaux assureront à la fois la photosynthèse mais porteront aussi les sporanges. Ces rameaux latéraux préfigurent l’apparition de la vraie feuille ou mégaphylle qui se distingue théoriquement du rameau latéral caulinaire par la distinction anatomique (passage d’une anatomie à symétrie radiale à une anatomie à symétrie bilatérale) et la présence d’un limbe ou surface plane. Ainsi les organes latéraux bien différenciés des « cladoxylopsides » du Dévonien (voir chapitre sur les Monilophytes), avec leur anatomie caulinaire à symétrie radiale et l’absence de vrai limbe, ne sont pas considérés comme des mégaphylles. Bien que ce soit encore bien étudié et discuté, les paléobotanistes s’accordent à proposer que la vraie mégaphylle serait apparue au moins deux fois indépendamment chez les Euphyllophytes, une fois chez les premières fougères et une fois chez les premiers Spermatophytes, et dans les deux cas au Dévonien supérieur. Ainsi la mégaphylle ou fronde des fougères n’est pas homologue de la mégaphylle des Spermatophytes. Adèle Corvez et collaborateurs dans une revue sur le sujet publiée en 2012 dans les Comptes Rendus Palévol proposent à ce titre de nommer « mégafronde » la feuille des fougères pour la distinguer de la mégaphylle des plantes à graines.
Pour ce qui est du système racinaire, de grandes inconnues subsistent. Le système racinaire adventif et à ramification dichotome des Lycophytes n’est pas homologue et est donc apparu indépendamment du système racinaire primaire et à ramification endogène (ayant très vraisemblablement pour origine une tige spécialisée, et qui peut devenir secondairement adventif par avortement du système primaire dans de nombreux groupes) observé actuellement chez tous les Euphyllophytes. Par contre l’apparition de ce système racinaire primaire et son origine chez les Euphyllophytes sont encore inconnues ou reposent sur des hypothèses hautement spéculatives.

Créez votre propre site internet avec Webador