Evolution anatomique
Evolution anatomique
Des hypothèses sur l’évolution des stèles chez les Trachéophytes peuvent être proposées. Les premiers Polysporangiophytes (POL) ont des analogues d’hydroides et leptoïdes avec une organisation et localisation qui préfigurent la protostèle circulaire ou haplostèle des premiers Trachéophytes du Silurien supérieur (TRA), telle qu’on l’observe chez les « rhyniopsides » fossiles. Cette protostèle se serait lobée pour donner indépendamment l’actinostèle exarche des Lycophytes (LYC) dès le Silurien supérieur et la protostèle tétra-lobée à protoxylème centrarche (les plus vieux éléments se forment au centre) des premiers Euphyllophytes au Dévonien inférieur. Chez les Lycophytes l’actinostèle donnera par fragmentation la plectostèle. Chez les Euphyllophytes, la protostèle tétra-lobée sera conservée chez les premiers Lignophytes du Dévonien moyen (LIG) et la migration du protoxylème donnera la protostèle tétra-lobée à protoxylème mésarche (confiné au centre des lobes) chez les ancêtres des Monilophytes du Dévonien inférieur (MON). Au sein des Monilophytes, la fragmentation de la protostèle donnera la plectostèle particulière des « cladoxylopsides » (CLA) au Dévonien moyen (chaque massif de tissus conducteurs s’entourant de bois) et vraisemblablement indépendamment la dictyostèle des fougères au Dévonien supérieur (FIL). L’inclusion des prêles (EQU) dans les fougères (voir chapitre correspondant) suggère que l’équisétostèle des Equisetidae dériverait d’une anatomie de fougère (nous y reviendrons plus tard). En parallèle, chez les Lignophytes, un cambium bifacial qui s’intercale entre le xylème et le phloème produit des tissus secondaires (bois et liber). L’eustèle des Spermatophytes (SPE) dériverait de la protostèle lobée par fragmentation. Le cambium peut être ou non fonctionnel, ce qui distingue les formes ligneuses des formes herbacées. Chez les Lilianae ou Monocotylédones (LIL), le cambium n’est plus fonctionnel et il y a prolifération des faisceaux sur plusieurs cercles pour donner l’atactostèle.
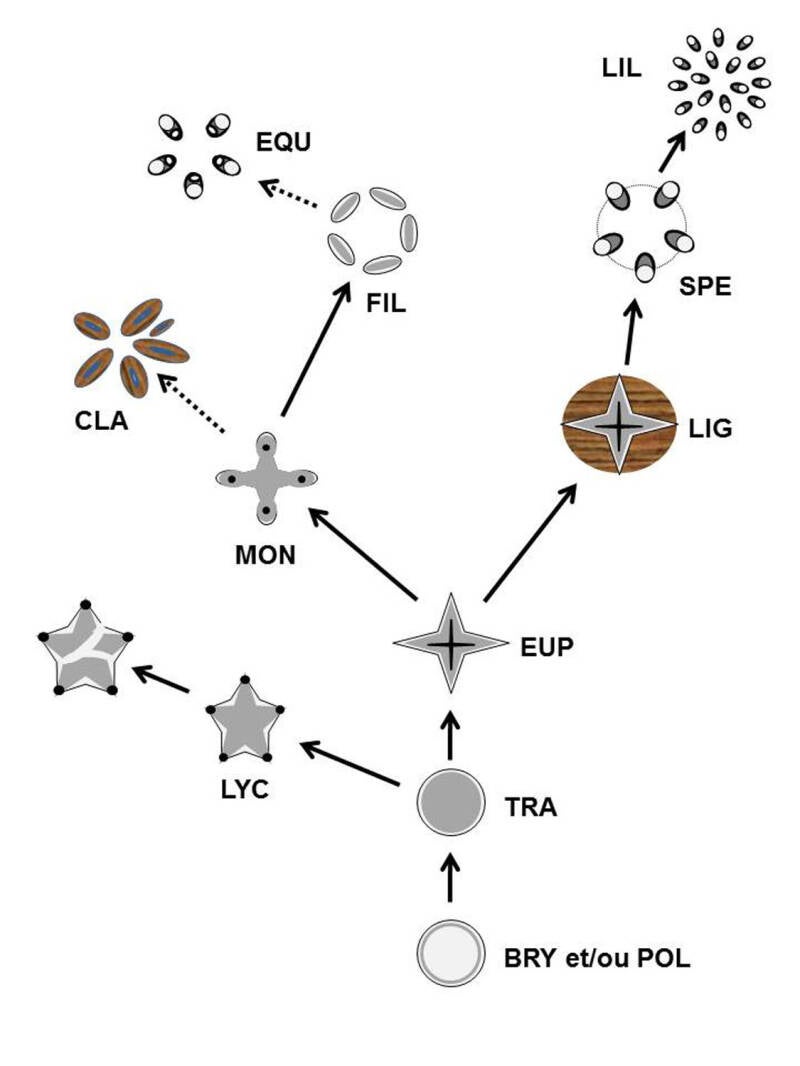
Evolution hypothétique de l’anatomie et des stèles (représentation schématique) chez les Embryophytes (détail dans le texte).
Le gris foncé représente le xylème, le gris clair le phloème (sauf chez premiers Polysporangiophytes, POL, où on a des analogues d’hydroïdes au centre et de leptoïdes à la périphérie), le noir la localisation du protoxylème (si utile), le fond texturé représente du bois, le cercle fin de l’eustèle des spermatophytes (SPE) la localisation du cambium qui peut ou ne pas être fonctionnel. Les flèches en pointillé indiquent une hypothèse sur des relations évolutives encore fortement discutées.
Chez les fougères, en plus du cas particulier de l’actinostèle des psilotes (Psilotidae), on observe une très grande diversité de stèles en dehors de la dictyostèle type. On trouve des protostèles types et des protostèles dites médullarisées (présentant un massif de parenchyme – un tissu fondamental de grosses cellules faiblement différenciées – au centre). Certaines fougères sont siphonostéliques, la stèle appelée siphonostèle (du mot siphon) est un cylindre creux avec du phloème des deux côtés (interne et externe), qui s’interrompt seulement au niveau des brèches foliaires, ces dernières ne produisant donc pas plusieurs méristèles en coupe transversale. Le phloème est majoritairement double, la siphonostèle est dite amphiploïque, mais il existe des cas minoritaires où le phloème interne est absent par perte secondaire, la siphonostèle est alors dite ectophloïque. Les siphonostèles ectophloïques peuvent être confondues avec les protostèles médullarisées mais seules les siphonostèles présentent des brèches foliaires. On observe des structures mixtes, une protostèle centrale entourée par une siphonostèle, une siphonostèle centrale entourée par une dictyostèle, ou d’autres variations. La polystélie, en particulier la poly-dictyostélie, n’est pas rare et se traduit par la superposition concentrique de plusieurs stèles d’un même type au sein d’une tige. La dictyostèle s’inscrit en fait avec la protostèle et la siphonostèle dans une série ontogénétique dont elle est le stade mature. Toute fougère quel que soit le type observé chez les individus matures passe par un stade protostélique chez le tout jeune sporophyte qui se creuse pour donner une siphonostèle, puis une dictyostèle par multiplication des brèches. Ainsi le cylindre plein donne un cylindre creux puis un réseau réticulé. Dans les eustèles (tout au moins chez les Spermatophytes) présentes dès les stades juvéniles, les faisceaux restent indépendants et déconnectés, il n’y a pas de réticulation et pas de vraies brèches foliaires. Le maintien chez certaines fougères de types anatomiques juvéniles dans les sporophytes matures illustrerait un phénomène de néoténie. Si on fait dériver l’équisétostèle des prêles d’une anatomie de fougère, le scénario le plus parcimonieux (qui prend en compte le moins de changements évolutifs) suggère que l’équisétostèle pourrait dériver de la fragmentation d’une siphonostèle ectophloïque (le phloème unique étant restreint du côté externe, donc vers la périphérie de la tige). Certaines phylogénies proposent une proximité évolutive entre Equisetidae et Ophioglossidae, ces dernières présentant des stèles ectophloïques, cela renforcerait l’hypothèse de l’évolution anatomique proposée.
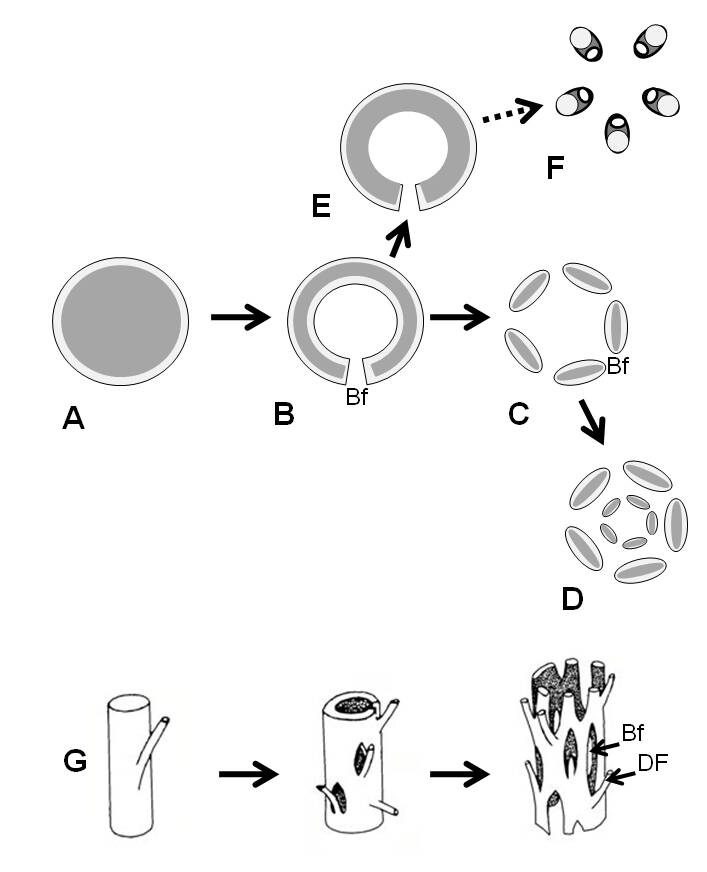
Diversité et évolution de l’anatomie (représentation schématique) chez les fougères (détail dans le texte).
Le gris foncé représente le xylème, le gris clair le phloème ;
A. Protostèle,
B. Siphonostèle amphiphloïque,
C. Dictyostèle,
D. Poly-dictyostèle,
E. Siphonostèle ectophloïque,
F. Equisétostèle des prêles,
G. même série que A-B-C mais en reconstruction 3D ; Bf = brèche foliaire, DF = départ de fronde.
Anatomie foliaire
La feuille dérivant de la tige, l’anatomie foliaire est elle-même dérivée de l’anatomie caulinaire. Elle s’en distingue par la symétrie, l’anatomie foliaire (tout au moins au niveau du pétiole) est à symétrie bilatérale alors que la symétrie caulinaire est radiale. On retrouve en effet cette symétrie bilatérale chez les microphylles de Lycophytes (à une seule nervure), les mégafrondes de fougères et les mégaphylles de Spermatophytes, trois organes apparus indépendamment (voir chapitre correspondant).
Anatomie racinaire
On s’accorde à proposer que l’anatomie des racines dérive également de l’anatomie caulinaire, tout en restant à symétrie radiale. Il existe une exception chez les Isoëtales (Lycophytes) avec une anatomie racinaire présentant une protostèle légèrement aplatie bilatéralement qui pourrait aller dans le sens d’une hypothèse que les racines des Isoëtales (et des Lepidodendrales éteintes apparentées) pourraient être des microphylles transformées. Chez les Euphyllophytes actuels, la racine présente une anatomie type actinostèlique à protoxylème exarche (on parle chez les Spermatophytes de différenciation centripète pour opposer la racine à la tige où la différenciation est centrifuge) et des massifs de phloème alternant avec les pôles de protoxylème. Chez les fougères, le xylème est réduit à deux pôles et la stèle d’une racine est alors semblable à une méristèle d’une dictyostèle caulinaire. Les racines actuelles des Spermatophytes sont intéressantes à étudier, le système vasculaire localisé au centre en actinostèle est bien délimité extérieurement par au moins une assise de cellules non différenciées à propriétés embryonnaires appelé péricycle. Au contact du péricycle et délimitant la partie corticale entourant la stèle, on observe une assise de cellules appelée endoderme à parois radiales épaissies et qui « contrôle » la diffusion des sèves de la stèle vers le cortex et réciproquement. Péricycles et endodermes ne se distinguent pas ou mal dans les organes caulinaires et foliaires des Spermatophytes (sauf quelques exceptions), mais sont par contre bien identifiables dans tous les organes chez les Trachéophytes non Spermatophytes en délimitant les tissus conducteurs quelle que soit leur organisation et donc le type de stèle (dans la dictyostèle par exemple, chaque méristèle est entourée par un péricycle puis un endoderme). Le péricycle est en fait encore présent dans les tiges des Spermatophytes, autour des faisceaux conducteurs, même si on le distingue mal, car il est à l’origine, via ses propriétés embryonnaires, des racines adventives. L’endoderme est visible au niveau caulinaire chez de nombreuses Angiospermes aquatiques, telles les Hydrocharitaceae avec les élodées, et dans les feuilles des Conifères. Son absence au niveau caulinaire et foliaire chez la majorité des Spermatophytes est due à une perte ou régression secondaire (ou plutôt, il peut potentiellement se différencier, mais cette différenciation ne s’exprime pas, plus ou partiellement). On notera que la racine des Spermatophytes avec son actinostèle et un endoderme bien différencié, illustrant une anatomie « archaïque », pourrait avoir conservé l’anatomie caulinaire des ancêtres communs.

Créez votre propre site internet avec Webador